Article Text

Abstract
Background: Because oxidative stress affects muscle function, the underlying mechanism to explain exercise induced peripheral muscle oxidative stress in patients with chronic obstructive pulmonary disease (COPD) is clinically relevant. This study investigated whether chronic hypoxaemia in COPD worsens peripheral muscle oxidative stress and whether an abnormal muscle inflammatory process is associated with it.
Methods: Nine chronically hypoxaemic and nine non-hypoxaemic patients performed repeated knee extensions until exhaustion. Biopsy specimens were taken from the vastus lateralis muscle before and 48 hours after exercise. Muscle oxidative stress was evaluated by lipid peroxidation (lipofuscin and thiobarbituric acid reactive substances (TBARs)) and oxidised proteins. Inflammation was evaluated by quantifying muscle neutrophil and tumour necrosis factor (TNF)-α levels.
Results: When both groups were taken together, arterial oxygen pressure was positively correlated with quadriceps endurance time (n = 18, r = 0.57; p<0.05). At rest, quadriceps lipofuscin inclusions were significantly greater in hypoxaemic patients than in non-hypoxaemic patients (2.9 (0.2) v 2.0 (0.3) inclusions/fibre; p<0.05). Exercise induced a greater increase in muscle TBARs and oxidised proteins in hypoxaemic patients than in non-hypoxaemic patients (40.6 (9.1)% v 10.1 (5.8)% and 51.2 (11.9)% v 3.7 (12.2)%, respectively, both p = 0.01). Neutrophil levels were significantly higher in hypoxaemic patients than in non-hypoxaemic patients (53.1 (11.6) v 21.5 (11.2) counts per fibre × 10−3; p<0.05). Exercise did not alter muscle neutrophil levels in either group. Muscle TNF-α was not detected at baseline or after exercise.
Conclusion: Chronic hypoxaemia was associated with lower quadriceps endurance time and worsened muscle oxidative stress at rest and after exercise. Increased muscle neutrophil levels could be a source of the increased baseline oxidative damage. The involvement of a muscle inflammatory process in the exercise induced oxidative stress of patients with COPD remains to be shown.
- FEV1, forced expiratory volume in 1 second
- FVC, forced vital capacity
- GPx, glutathione peroxidase
- HR, heart rate
- MCTSACT, mid thigh muscle cross sectional area
- MVC, maximal voluntary contraction
- Pao2, arterial oxygen pressure
- ROS, reactive oxygen species
- SOD, superoxide dismutase
- Spo2, oxygen pulse saturation
- TBARs, thiobarbituric acid reactive substances
- TNF-α, tumour necrosis factor α
- chronic obstructive pulmonary disease
- exercise
- hypoxaemia
- lipid peroxidation
- lipofuscin
- oxidised protein
- inflammation
Statistics from Altmetric.com
- FEV1, forced expiratory volume in 1 second
- FVC, forced vital capacity
- GPx, glutathione peroxidase
- HR, heart rate
- MCTSACT, mid thigh muscle cross sectional area
- MVC, maximal voluntary contraction
- Pao2, arterial oxygen pressure
- ROS, reactive oxygen species
- SOD, superoxide dismutase
- Spo2, oxygen pulse saturation
- TBARs, thiobarbituric acid reactive substances
- TNF-α, tumour necrosis factor α
- chronic obstructive pulmonary disease
- exercise
- hypoxaemia
- lipid peroxidation
- lipofuscin
- oxidised protein
- inflammation
Exercise induced oxidative stress has been documented in the pulmonary, blood, and muscle compartments of patients with chronic obstructive pulmonary disease (COPD).1–3 Because oxidative stress can affect skeletal muscle contractibility and function,4 it has been proposed as a potential mechanism of peripheral muscle dysfunction and impaired exercise tolerance in this disease.5 Indeed, oral antioxidant supplementation with N-acetylcysteine improves quadriceps endurance in COPD,6 supporting a role for exercise induced oxidative stress in diminishing quadriceps endurance in these individuals.
Oxidative stress is considered to be present when the highly reactive oxygen species (ROS) overwhelm the antioxidant defences. The role of hypoxia in modulating oxidative metabolism and production of reactive oxygen derived metabolites has been investigated in several in vitro and animal studies.7,8 Although these experiments have shown that the source of ROS production in hypoxia is likely to be the mitochondria, particularly the respiratory chain,8,9 the current understanding of the effects of hypoxia on in vivo human oxidative stress is only fragmentary.
The available studies have focused on long term exposure to hypoxic conditions in healthy subjects. In these individuals, oxygen deficient environmental conditions such as those encountered during high altitude expeditions are associated with increased ROS production and oxidative stress in the systemic circulation at rest and during exercise.10,11 Little is known regarding the effects of hypoxaemia on skeletal muscle oxidative stress in humans. Two studies have reported cumulative oxidative stress and damage in resting muscle of healthy subjects exposed to an oxygen deficient environment. Martinelli and co-workers12 reported a threefold increase in basal levels of lipofuscin, a pigment marker of cumulative oxidative stress, in the vastus lateralis of climbers after a high altitude expedition over 5000 m for 8 weeks. More recently, Lundby and colleagues13 reported an increase in resting muscle oxidative DNA damage in seven healthy subjects after 2 weeks at 4100 m. Chronic hypoxaemia, a common physiopathological consequence of COPD, is associated with poor tolerance of peripheral muscle exercise.14 The literature on environmental hypoxia suggests that chronic hypoxaemia in COPD could contribute to peripheral muscle oxidative stress under resting conditions and after exercise.12,13 The primary objective of this study was therefore to test whether chronic hypoxaemia worsens peripheral muscle oxidative stress in patients with COPD at rest and in response to exercise.
The underlying mechanisms for hypoxia induced oxidative stress have yet to be investigated in humans. In vitro studies suggest that inflammation, via the release of cytokines or neutrophils, is one potential pathway.15 In cultured cells, hypoxia resulted in an activation of nuclear factor kappa-B, a central transcription factor involved in the orchestration of inflammatory cascade events.16,17 In COPD, reduced lung function is associated with increased blood inflammatory markers.18 Furthermore, Takabatake and co-workers showed that arterial oxygen tension (Pao2) is inversely correlated with circulating tumour necrosis factor alpha (TNF-α) and its soluble receptor,19 suggesting that hypoxaemia in COPD could be associated with the development of a systemic inflammatory process. These results led to the hypothesis that hypoxaemia could worsen muscle oxidative stress through the induction of an inflammatory process in the peripheral skeletal muscles. The second objective of this study was therefore to investigate whether an abnormal inflammatory process occurs at rest and after exercise in the peripheral muscles of hypoxaemic COPD patients.
To achieve these two objectives, we investigated the effect of chronic hypoxaemia on oxidative stress and inflammatory mediators in the vastus lateralis muscle of COPD patients before and after localised muscle exercise designed to minimise the demand on the cardiorespiratory system.14,20
METHODS
Subjects
Eighteen ex-smoking men with stable severe COPD as defined by the GOLD guidelines21 volunteered to participate in the study. Patients were categorised into two groups according to their resting Pao2 whose cut off value corresponded to the lower normal limit as predicted by the Mellemgaard reference equation.22 One group consisted of non-hypoxaemic patients (n = 9, Pao2 >lower normal limit) and the other consisted of chronic hypoxaemic patients (n = 9, Pao2 <lower normal limit). The exclusion criteria were unstable disease (that is, COPD exacerbation in the preceding 2 months), long term supplemental oxygen, sleep related breathing disorders, and other potentially confounding factors such as neuromuscular disease, chronic heart failure, diabetes mellitus, or alcoholism. The pharmacological treatment for all patients included inhaled β agonists and/or anticholinergics, and in four cases inhaled corticosteroids. No patients were on theophylline or oral steroid therapy either at study entry or in the preceding 6 months. Patients were considered as sedentary (scores <9) according to a physical activity questionnaire adapted for older and retired subjects and used for COPD patients.3 All participants had a body mass index lower than 30 and all were questioned on their dietary habits to rule out antioxidant or vitamin supplementation. Informed written consent was obtained from all patients and the research protocol was approved by the local institutional ethic committee.
Study design
Patients were instructed to abstain from strenuous physical activity for 4 days before and 2 days after being studied. On day 1 the subjects underwent spirometric testing, arterial blood sampling, and computed tomographic (CT) scanning of the thigh, and they had to answer the physical activity questionnaire. Strength was evaluated in each leg and a baseline muscle biopsy was performed on the non-dominant leg. On day 2 the subjects were familiarised with the endurance test procedures by performing five consecutive dynamic knee extensions of the dominant leg. They then performed the localised muscle endurance exercise test. On day 4 (48 hours after localised exercise) a biopsy specimen of the dominant quadriceps was obtained.
Study methods
Pulmonary function tests and blood gas analysis
All subjects underwent spirometric testing from which forced expiratory volume in 1 second (FEV1) and forced vital capacity (FVC) were obtained and the FEV1/FVC ratio calculated. The results of pulmonary function testing were compared with the normal values of Knudson and co-workers.23 Arterial blood was drawn from a radial artery while the subject was breathing room air in the seated position. Pao2 was then analysed with a blood gas analyser (AVL 995, AVL Scientific, Roswell, GA, USA).
Mid thigh muscle cross sectional area measurement
A CT scan of the dominant thigh (that is, the stronger one) was performed halfway between the pubic symphysis and the inferior condyle of the femur using a fourth generation Toshiba Scanner 900S (Toshiba Inc, Tokyo, Japan). The mid thigh muscle cross sectional area (MTCSACT) was obtained as previously described.24
Quadriceps strength measurement
The maximal voluntary strength of the quadriceps was measured in a standardised seated position while the subjects performed dynamic knee extensions against a hydraulic resistance (HF STAR, Hydrafitness Total Power; Henley Health Care, Belton, TX, USA) as previously described.3
Quadriceps endurance measurement
Quadriceps endurance was assessed on an exercise bench according to the technique described by Couillard et al.3 The exercises were performed with the subject in a seated position at 90° knee and hip flexion with arms crossed on the chest, and the position maintained to avoid any changes in respiratory responses. The subjects were asked to perform repeated knee extensions of the dominant leg against weights corresponding to 30% of their maximal voluntary contraction, at a pace of six movements per minute imposed by an audio signal (metronome) until exhaustion. The dynamic knee extension was performed for 3 seconds, immediately followed by active leg return (eccentric flexion) against resistance for 3 seconds and by rest (for 4 seconds) before the next extension. The test was stopped when the subjects could no longer perform a maximal knee extension or maintain the imposed rhythm despite verbal encouragement. The duration of the test was then recorded as the quadriceps endurance time. The sensations of dyspnoea and muscle fatigue were measured at rest and immediately after exercise on a 10 cm visual analogue scale. Oxygen pulse saturation (Spo2) and heart rate were measured throughout both exercise protocols using a pulse oximeter (Ohmeda Biox II 3740 Pulse Oximeter).
Skeletal muscle analysis
Muscle biopsies and tissue preparation of the vastus lateralis
Percutaneous biopsies of the vastus lateralis muscle (200–300 mg) of the non-dominant leg at baseline and of the dominant leg 48 hours after exercise were performed 10 cm above the patella as described by Bergström.25 The samples for measurement of oxidative stress markers were immediately frozen in liquid nitrogen and then stored at −80°C until processing. For histochemical analysis, the samples were embedded in OCT compound (Tissue-Tek, Miles Inc, Elkhart, IN, USA) and then stored at −80°C until transverse sections were cut using a cryostat Leica Jung CM 3000 (Wetzlar, Germany) at −20°C. Each section was verified by light microscopy to ensure proper fibre orientation. All muscle specimens were coded and analysed without knowledge of the subject’s clinical state.
Fibre type determination
Histochemical fibre typing of the biopsy specimens was carried out as described previously.26 Briefly, muscle samples were cut into 10 µm thick transverse sections in a cryostat at −20°C and stained for myofibrillar adenosine triphosphatase activity, according to the single step ethanol modified technique.27 Staining intensity designated fibre types as type I (non-stained), type IIa (lightly stained), and type IIx (darkly stained). The mean (SE) total number of fibres counted for muscle fibre typing was 405 (140) (95% CI 82 to 728) and 301 (48) (95% CI 290 to 412) for the hypoxaemic and non-hypoxaemic groups, respectively.
Determination of muscle oxidative stress markers
Muscle lipofuscin inclusion accumulation, a marker of cumulative oxidative stress, was measured only in pre-exercise biopsy specimens according to the method previously described by Allaire and co-workers.28 Thiobarbituric acid reactive substances (TBARs) were used as markers of muscle lipid peroxidation and were determined spectrophotometrically using the method described by Ohkawa and co-workers.29 The final results were expressed in nmol/g wet weight. The reproducibility of TBARs, which was calculated as coefficients of variation, was less than 10%.3 In the subjects in whom a sufficient amount of muscle tissue was still available (control group, n = 7; hypoxaemic group, n = 8), protein oxidation was also measured by evaluating the levels of protein carbonyls using immunoblotting (Oxyblot kit; Serological Corporation, Norcross, GA, USA). Muscle protein carbonyl contents were calculated by adding the integrated density of individual protein bands (Alpha Innotech Corporation, San Leandro, CA, USA) obtained by Western blot analysis.3
Determination of muscle antioxidant activity
The muscle activity of glutathione peroxidase (GPx) and superoxide dismutase (SOD) was quantified spectrophotometrically according to the methods of Nakamura et al30 and Sun et al,31 respectively. A unit of SOD activity is defined as the amount of SOD required to cause 50% inhibition of the oxidation of epinephrine. The reproducibility of these measurements, which were calculated as coefficients of variations, was less than 10%.
Determination of muscle inflammatory mediators
The OCT embedded frozen muscle tissue was cut into 10 µm sections, mounted on slides, and processed for immunohistochemical analysis. Serial sections were used for immunostaining of TNF-α and neutrophils using human specific antibodies. The tissue sections were fixed at −20°C in acetone/methanol and rinsed in phosphate buffered saline (PBS). To block the non-specific binding of antibodies to tissue, slides were incubated in a humid chamber with a peroxidase blocking reagent 0.3% for 30 minutes at room temperature (DAKO Diagnostics Canada Inc, Mississauga, ON, Canada), rinsed with PBS, and then preincubated with a universal blocker (DAKO). Subsequently, sections were incubated at 4°C for 2 hours in the humid chamber with monoclonal anti-human TNF-α (Biosource International, Camarillo, CA, USA; dilution 1/25) or monoclonal anti-human elastase (DAKO; dilution 1/200) primary antibodies. After several washes in PBS, the slides were incubated for 1 hour with horseradish peroxidase secondary antibodies (anti-mouse Ig horseradish peroxidase-link whole antibody, Amersham Biosciences Corporation, Piscataway, NJ, USA; dilution 1/100 in PBS). The sections were then stained using amino ethyl carbazol (AEC) and chromogen as a substrate (DAKO). Slides were counterstained with Mayer’s haematoxylin. Positive controls consisted of bronchoalveloar lavage cells from patients with asthma. For negative controls the primary antibody was omitted.
Image analysis and calculations
Muscle sections were magnified and transmitted to image analysis software (Image Pro Plus 4.1 for Windows; MediaCybernetics, Silver Springs, MD, USA). For each subject the fibre type composition was calculated as the total number of fibres of a given type divided by the total numbers of fibres. The mean fibre size was calculated based on the cross sectional area on 150 randomly selected fibres.26 Lipofuscin inclusions were defined as dark blue dots in the pale blue green muscle cell cytosol (fig 1A). Quantification of lipofuscin inclusions was performed at a magnification of 200× on approximately 150 random fibres. The results are expressed as the lipofuscin/fibre ratio. The muscle inflammatory mediators, as determined by semi-quantitative analyses, are expressed as the total number of positive fibres divided by the total number of fibres (fig 1B, C). All sections were analysed in a blinded fashion and in random order by two observers who were unaware of the clinical data of the case under study. The agreement between the observers was assessed by plotting individual differences between observers’ counts against their respective means, as described by Bland and Altman (see fig E1 in online supplement).32

Representative immunohistochemical specificity staining. (A) Lipofuscin inclusions were easily identified as they appeared as dark blue dots in the pale blue green muscle cell cytosol from patients with COPD. Magnification ×200. (B) TNF-α immunopositive staining in bronchoalveolar lavage cells from a patient with asthma. Magnification ×20. (C) Neutrophil immunopositive staining in the muscle cell cytosol from patients with COPD. Magnification ×100.
Statistical analysis
Values are reported as mean (SE) with 95% confidence intervals (CI). Clinical data and the endurance time were compared between the two groups of COPD patients using an unpaired t test or a Mann-Whitney rank sum test when normality was not obtained. When a t test was performed to compare group mean difference, 95% CI and p values are noted. When a Mann-Whitney rank sum test based on group median difference was used, only a p value is reported. A two way analysis of variance followed by Tukey’s pairwise multiple comparison procedure was used to determine the effects of hypoxaemic status and localised exercise on the biological markers. The significance level was set at 0.05. The data were analysed using the statistical package Sygmastat 1.0 (Jandel Scientific, San Rafael, CA, USA).
RESULTS
Anthropometric and spirometric data
There were no differences in the anthropometric and spirometric data between the two groups (table 1). On average, patients showed severe to very severe airflow obstruction with a mean (SE) FEV1 of 35 (3)% (95% CI 29 to 41) of predicted values.23
Anthropometric and spirometric data
The mean (SE) predicted Pao2 for all patients was 11.34 (0.04) kPa (95% CI 11.25 to 11.43), with a lower normal value of 9.74 (0.04) kPa (95% CI 9.65 to 9.83). The mean Pao2 averaged 8.69 (0.20) kPa (95% CI 8.23 to 9.15) and 10.41 (0.26) kPa (95% CI 9.81 to 11.01) for the hypoxaemic and non-hypoxaemic groups, respectively (mean difference −1.72 (95% CI 1.02 to 2.41); p = 0.0001). This represented 76 (2)% (95% CI 72 to 80) and 92 (2)% (95% CI 87 to 97) of the predicted value, respectively (mean difference −15 (95% CI 10 to 21); p = 0.0001).
Muscle function and local muscle exercise parameters
Mid thigh muscle cross sectional area and maximal voluntary contraction were comparable between the two groups (table 2). Localised muscle endurance tests were performed at a mean load corresponding to 9.0 (0.5) kg (95% CI 7.7 to 10.3). The hypoxaemic group had similar quadriceps endurance time to that of the non-hypoxaemic group (table 2) but, when both groups were taken together, a significant and positive correlation was noted between resting Pao2 and quadriceps endurance time (fig 2). The exercise induced fall in oxygen pulse saturation (Spo2) was significantly larger in the hypoxaemic group than in the non-hypoxaemic group (table 2). Indeed, Spo2 remained constant throughout exercise in the non-hypoxaemic group while a significant fall in Spo2 was observed in the hypoxaemic group (table 2). The exercise induced increase in heart rate was of small amplitude. The increase in the muscle fatigue score on the visual analogue scale during localised muscle exercise was greater than that of dyspnoea (mean difference −3.1 (95% CI −4.4 to −1.7); p = 0.0001).
Muscle function and local muscle exercise parameters

Positive correlation between Pao2 (kPa) and quadriceps endurance time (s) during localised exercise in the 18 patients with COPD (solid symbols represent hypoxaemic patients and open symbols represent non-hypoxaemic patients; p = 0.02, r = 0.57).
Skeletal muscle data
Fibre type composition and mean fibre size
The fibre type distribution was characterised by a decrease in type I fibres (32 (4)% (95% CI 25 to 39)) and a corresponding increase in type II fibres, with no significant difference between groups (table 3). Mean fibre size was also identical between the two groups.
Fibre type distribution and fibre size in the non-hypoxaemic and hypoxaemic groups
Oxidative stress markers
More than 90% of the lipofuscin inclusions observed in our patients were located in the subsarcolemmal area (fig 1A). The number of lipofuscin inclusions per fibre in pre-exercise muscle samples was significantly greater in the hypoxaemic group than in the non-hypoxaemic group (2.9 (0.2) v 2.0 (0.3) lipofuscin inclusions/fibre, respectively; mean difference +0.9 (95% CI +0.1 to +1.8); p = 0.032). No significant difference was observed in baseline TBARs (142.4 (119.0) v 155.2 (24.2) nmol/g wet weight; mean difference −12.8 (95% CI −70.0 to +44.5), p = 0.64) and protein carbonyl contents (15.3 (4.1) v 16.2 (3.9) integrated density ×104; mean difference −0.9 (95% CI −9.6 to +7.8), p = 0.82) between the hypoxaemic and non-hypoxaemic groups.
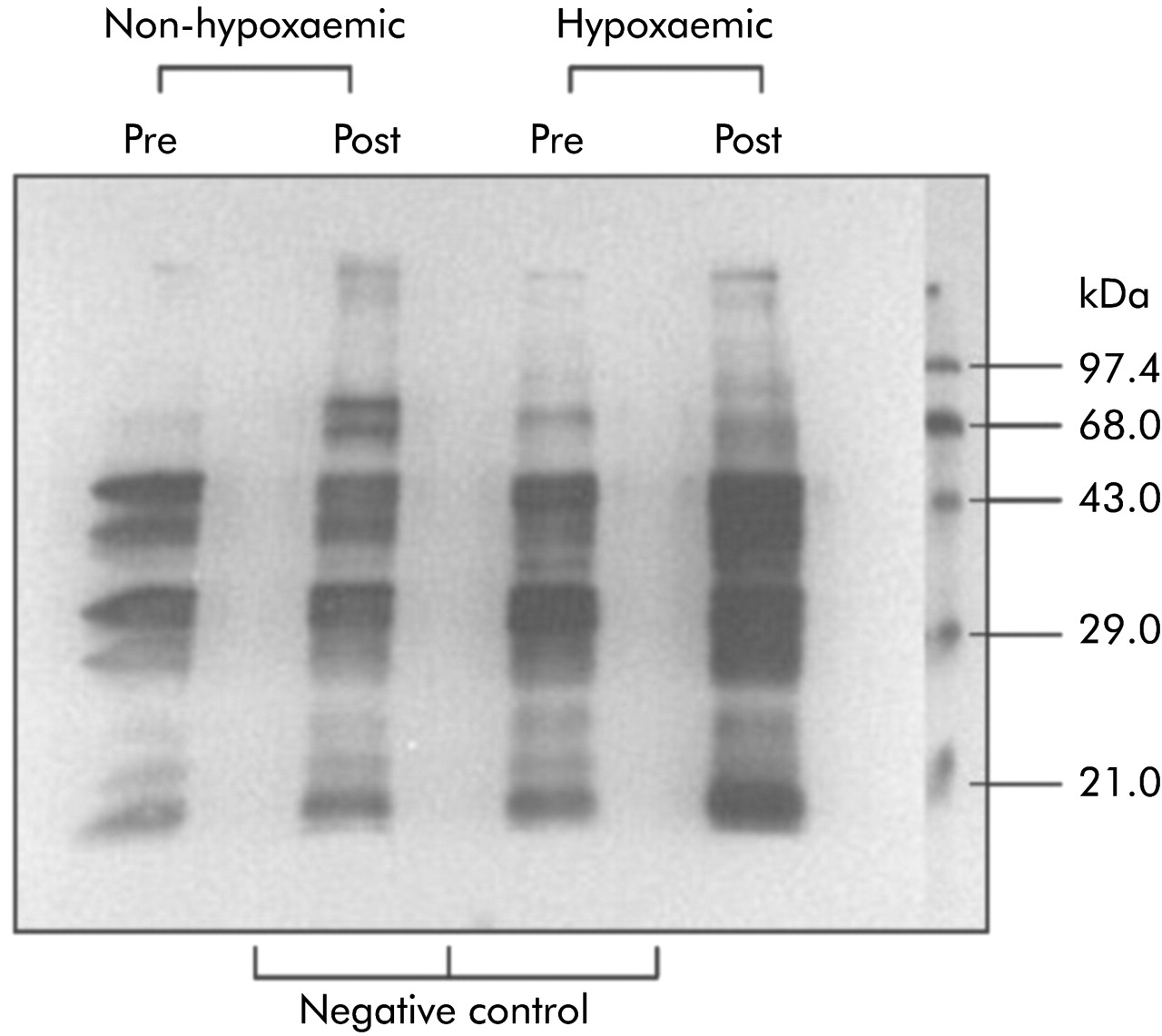
Representative immunoblots of the muscle protein carbonyl group obtained in a hypoxaemic and a non-hypoxaemic patient before and after exercise are shown in fig 3. Four to five bands with molecular weights varying from 20 kDa to 68 kDa, including two main 40 and 30 kDa protein bands, were detected in the COPD patients. The baseline migration pattern of these bands was similar between the two groups. After quadriceps exercise these bands became more intense only in the hypoxaemic patients, but no new bands were detected.

Representative immunoblots of muscle protein carbonyl groups in a non-hypoxaemic patient and a hypoxaemic patient before (pre) and after (post) quadriceps exercise. Four to five bands whose molecular weight varied from 20 to 68 kDa were detected in both patients. Compared with the pre-exercise levels, these bands became more intense in the hypoxaemic patient but no new bands were detected. Negative controls (underivatised protein) are also shown.
The group mean change in muscle TBARs and oxidised proteins 48 hours after exercise are shown in fig 4 for the two groups of patients. There was a greater increase in these oxidative stress markers in the hypoxaemic group than in the non-hypoxaemic group (p = 0.01).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of localised muscle exercise on the increase in muscle lipid peroxidation and oxidised protein levels in the non-hypoxaemic group (open bars; n = 9) and the hypoxaemic group (black bars; n = 9). Both markers of oxidative stress increased markedly after exercise, with a significantly greater increase in the hypoxaemic patients than in the non-hypoxaemic patient (40.6 (9.1)% v 10.1 (5.8)%; mean difference +30.4 (95% CI +7.6 to +53.3), p = 0.01 for TBARs; 51.2 (11.9)% v 3.7 (12.2)%; mean difference +47.5 (+10.6 to +84.4); p = 0.01 for oxidised protein). *p<0.05 hypoxaemic v non-hypoxaemic group.
Enzymatic antioxidant activity
Similar baseline muscle GPx and SOD activities were observed in the two COPD groups (table 4). In both groups, exercise failed to induce any increase in GPx or SOD activities.
Muscle enzymatic antioxidant activity and neutrophil counts at rest and after exercise in the non-hypoxaemic and hypoxaemic groups
Inflammatory markers
The number of neutrophil counts per muscle fibre was higher in the hypoxaemic group than in the non-hypoxaemic group (group effect: 53.1 (11.6) v 21.5 (11.2) counts per fibre × 10−3; p = 0.04). Exercise did not alter the muscle neutrophil levels in either group (table 4). Muscle TNF-α was not detected by immunostaining in the biopsy specimens taken at rest or after exercise in either group of patients.
DISCUSSION
The major findings of this study are that hypoxaemia worsened muscle oxidative stress in COPD at rest and after localised muscle exercise. Increased muscle neutrophil levels were observed at rest in the muscle biopsy specimens from hypoxaemic patients, suggesting a greater level of muscle inflammation in these individuals. TNF-α was not detected in the biopsy specimens taken either at rest or after exercise.
Chronic hypoxaemia and peripheral muscle oxidative stress in COPD patients
Evidence of lipofuscin accumulation, and thus cumulative lipid peroxidation, has been reported in the resting skeletal muscle of patients with COPD.28 This study adds to the current knowledge by showing that COPD patients with decreased resting Pao2 have a greater lipofuscin accumulation in the vastus lateralis muscle than patients with normal Pao2. This observation could not be explained by differences in muscle fibre size or composition between the non-hypoxaemic and hypoxaemic patients. The greater muscle accumulation of lipofuscin observed in the hypoxaemic group thus supports the hypothesis that hypoxaemia, as a chronic condition, worsens basal cumulative oxidative damage in COPD skeletal muscle. After exercise the significant increase in muscle TBARs and protein carbonyls (two markers of acute oxidative stress) indicated quadriceps exercise induced oxidative stress,3 particularly in the hypoxaemic patients whose muscle TBAR and protein carbonyl levels after exercise were greater than in the non-hypoxaemic patients. These results are therefore consistent with the hypothesis that chronic hypoxaemia worsens peripheral muscle oxidative stress in COPD, both at rest and in response to exercise. The exact mechanisms by which chronic hypoxaemia worsens in vivo peripheral muscle oxidative stress at rest and in response to exercise remain unknown. Nevertheless, some hypotheses in line with our results can be proposed in COPD.
Potential underlying mechanisms
Similar changes in the activities of SOD and GPx were observed in the two groups, suggesting that differences in these enzymatic antioxidant defences did not account for the greater muscle oxidative stress observed in hypoxaemic patients. However, the increased neutrophil count observed in the quadriceps of hypoxaemic patients may have contributed to the increased basal oxidative stress in these patients. Indeed, circulating neutrophils are important effector cells in the generation of free radicals via the reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase system.33,34 Neutrophils and phagocytes may thus be critical cells in the pathogenesis of hypoxaemia related oxidative stress, supporting the notion that inflammatory mediators are involved in the basal cumulative oxidative stress in the thigh muscle of hypoxaemic COPD patients.
However, no TNF-α could be detected in the resting or post-exercise muscle biopsy specimens of either group of patients. Similarly, no changes in muscle neutrophil levels were observed after exercise. These results may indicate that exercise, which was localised to one muscle group for only a few minutes, was not sufficient to induce an inflammatory process in the muscle of hypoxaemic and non-hypoxaemic COPD patients, despite evidence of exercise induced muscle oxidative stress. However, a role for a muscle inflammatory process in the exercise induced oxidative stress cannot be excluded from the present results. Indeed, a number of mediators other than neutrophils and TNF-α may be involved in the development of an inflammatory process within the muscle. For instance, an increase in nuclear factor NF-κB, a key inflammatory transcription factor,35 has recently been observed in the skeletal muscles of COPD patients with low body weight.36 Furthermore, a possible muscle inflammatory process could have been missed since the post-exercise muscle biopsy specimen was obtained at a single time point. Whether muscle inflammation was present in exercising muscle—and whether it played a role in the muscle oxidative stress—thus remains unclear and needs to be examined further.
Based on the location of lipofuscin pigments, which were mostly in the mitochondria-rich subsarcolemmal area, one possibility is that the mitochondrial electron transportation chain was responsible for the increased ROS production in the skeletal muscle of COPD patients with hypoxaemia. Evidence for increased mitochondrial ROS production has been observed during cellular hypoxia.8,9 The xanthine oxidase system in skeletal muscle is another important source of ROS production in conditions such as the ischaemia-reperfusion phenomenon which is associated with excessive production of free radicals and oxidative injuries.37 Based on the increase in blood flow in working muscles during exercise leading to muscle reoxygenation, it may be that exercise in hypoxaemic patients could mimic this ischaemia-reperfusion phenomenon. Given the role of the xanthine oxidase system in inducing oxidative stress during exercise in non-hypoxaemic patients with COPD,38 this system is also likely to contribute to the enhanced exercise induced oxidative stress in hypoxaemic patients with COPD.
Hypoxaemia induced muscle oxidative stress and peripheral muscle function
Evidence of increased molecular oxidative damage, reflected by increased lipid peroxidation and oxidised proteins, was found in the quadriceps of hypoxaemic patients at rest and in response to exercise. However, we did not document functional muscle changes as the strength and endurance of the quadriceps of the hypoxaemic and non-hypoxaemic patients were not statistically different. This may suggest that the molecular oxidative damage associated with the mild hypoxaemia in this study was not sufficient to alter muscle function. Findings from studies carried out at high altitude suggest that peripheral muscle strength is not affected by oxygen deficient environmental conditions.39,40 Although acute or chronic exposure to hypobaric hypoxia in healthy subjects impairs large muscle exercise performance, the effects of hypoxic conditions on the endurance of isolated small muscle groups are poorly defined.41 Nevertheless, some studies have shown reduced local muscle endurance under hypoxic conditions in healthy subjects and in patients with COPD.41,42 In the present study we found that quadriceps endurance in COPD patients was inversely correlated with the severity of hypoxaemia, as previously shown by Serres et al.13 Because higher muscle oxidative stress was observed in the hypoxaemic patients in our study, and oxidative stress has recently been shown to alter quadriceps muscle endurance in patients with COPD,6 we can hypothesise that peripheral muscle oxidative stress may be a deleterious in vivo mechanism through which hypoxaemia could lead to skeletal muscle dysfunction in COPD.2
Methodological considerations
Hypoxaemic status and analysis of patient subgroups
We specifically excluded from the study patients qualifying for long term oxygen therapy (Pao2 <7.3–8.0 kPa). In these patients it would be unethical to delay oxygen therapy for research purposes and we wished to avoid the potential confounding effects of supplemental oxygen on oxidative stress. We used the Mellemgaard reference equation usually used in clinical hospital practice to detect hypoxaemia, since it takes into account the age in determining the cut off value of predicted normal Pao2 values.22 The hypoxaemic group included patients with a Pao2 >8.0 kPa but lower than the lower normal limit based on the Mellemgaard predictive equation.22 Despite being only mildly hypoxaemic, their oxygenation status was clearly different from that of the non-hypoxaemic group (mean resting Pao2 1.72 (0.41) kPa lower than in the non-hypoxaemic group and, in contrast to the latter group, they showed a significant fall in Spo2 during exercise). Given that our two subgroups of patients were otherwise comparable, it is reasonable to assume that differences in blood oxygenation were involved in the observed difference in the peripheral oxidative stress and inflammatory status between them.
Quadriceps exercise protocol
The localised muscle exercise used in the study to assess muscle endurance has been discussed previously in terms of its reproducibility and specificity to the quadriceps.3 Previous electromyographic recordings of the thigh indicated that only the quadriceps was active during this localised exercise.20 The increases in dyspnoea sensation and heart rate following exercise were of small amplitude. In contrast, the increase in the perception of muscle fatigue was higher, suggesting that the main reason for stopping exercise was peripheral and not related to respiratory or cardiovascular limitations.
Markers of muscle oxidative stress
A useful feature of lipofuscin is its tendency to accumulate slowly over time, as it cannot be eliminated effectively.43 Since lipofuscin accumulation reflects the magnitude of lipid peroxidation, it can thus be used as a marker of cumulative oxidative stress and has therefore been evaluated only in pre-exercise biopsy specimens.28,43 In contrast, TBARs were used as a marker of oxidant mediated lipid damage induced by exercising muscle. One potential limitation of TBARs is that, under oxidative stress conditions, malondialdehyde, hydroperoxides, and some carbohydrates and amino acids may yield products that are able to react with thiobarbituric acid.44 However, the spectrophotometric method that we used was sufficiently sensitive and reproducible to provide a valid estimation of oxidative stress,29 as previously discussed.3 Protein carbonyls are sensitive indices of oxidative injury45 and the detection of muscle oxidised proteins provided another fingerprint of exercise oxidative damage and strong evidence of exercise induced oxidative stress in our patients.
Timing of the post-exercise biopsy
The timing of peak values for an exercise induced increase in muscle oxidative stress markers is a highly debatable issue and, according to previous studies, has been found to vary from immediately after exercise to 72 hours after exercise.46–49 In this study the biopsy specimens were obtained 48 hours after exercise, as in a previous study by our group which showed increased oxidative stress in the quadriceps of non-hypoxaemic COPD patients 48 hours after an exercise protocol similar to the one used here.3 Since the mitochondria respiratory chain is one of the major sources of oxidant production during exercise, one can predict that the highest rate of ROS production and increased muscle lipid peroxidation should occur immediately after the cessation of exercise. However, ROS were not directly measured; rather, indirect markers of their presence (TBARs and oxidised proteins) which could be detected for a much longer period of time were evaluated.46–49 The timing of the second biopsy thus appears appropriate in evaluating the exercise induced muscle inflammation response.50
Conclusion
This study shows that chronic hypoxaemia in COPD patients worsened peripheral muscle oxidative stress both at rest and during exercise. An increase in muscle neutrophil levels could be one potential mechanism of oxidative stress in skeletal muscle at rest. The presence and role of a muscle inflammatory process in the worsening of oxidative stress after exercise remain to be demonstrated. Understanding the underlying mechanisms by which hypoxaemia affects peripheral muscle oxidative stress in COPD will probably lead to the development of better therapeutic approaches for peripheral muscle dysfunction in this disease.
Acknowledgments
The authors thank Marie-Josée Breton, Marthe Bélanger, and Catherine Stott-Carmeni for valuable technical assistance.
REFERENCES
Supplementary materials
The figure is available as a downloadable PDF (printer friendly file).
If you do not have Adobe Reader installed on your computer,
you can download this free-of-charge, please Click hereFiles in this Data Supplement:
- [view PDF] - Figure E1. Plots of the differences for the two observers in neutrophil counts against their means, according to the Bland and Altman method[32] (n = 36).
Footnotes
-
Published Online First 17 June 2005
-
This study was supported in part by a Canadian Institutes of Health Research grant number MOP-53135
-
Competing interests: CK was supported by a travelling grant from La Cooperation Franco-Québécoise. FM is a research scholar of the Fonds de la Recherche en Santé du Québec. DS is a recipient of a PhD training award from the Fonds de la Recherche en Santé du Québec.